Why I think the Jurassic/Cretaceous boundary is super important
Mass extinctions are insanely catastrophic, but important, events that punctuate the history of life on Earth. The Jurassic/Cretaceous boundary, around 145 million years ago, was originally thought of to represent a mass extinction, but has subsequently been ‘down-graded’ to a minor extinction event based on new discoveries.
However, compared to other important stratigraphic boundaries, like the end-Triassic or the end-Cretaceous, both time periods representing mass extinction events, the Jurassic/Cretaceous (J/K) boundary actually remains really poorly understood. This is both in terms of what was going on with different animal groups at the time, and what environmental changes were occurring alongside this.
Well, I have a new research paper out now that synthesises more than 600 research articles, bringing them together to try and build a single picture of what was going on around this time! It’s free to read here, and is essentially the literature review from my thesis, or as I like to think of it, the justification for my existence as a researcher!
I don’t want to go into too many details here, but it provides detailed accounts of the environmental changes occurring across the J/K transition, and how this impacted upon different plant and animal groups throughout this period.
The main conclusions I think are worth highlighting here, so you don’t have to read the whole thing!
- The Late Jurassic–Early Cretaceous interval represents a time of major biotic upheaval and reorganisation. The precise magnitude of extinction is currently unknown, especially in light of our increasing awareness of the impact of incomplete sampling on the patterns preserved in the fossil record. However, it is clear that the J/K extinction, although severe in multiple groups, was not on the same scale as that for the ‘Big Five’ mass extinctions. What is becoming apparent, though, is that the J/K interval represents a period of elevated extinction, substantially protracted over some 25 million years, and involves the persistent loss of diverse lineages, and the origins of many major groups that survived until the present day (e.g. birds).
- There is widespread evidence for a major faunal turnover in both the marine and terrestrial realms during the J/K interval. Whereas the effect of this is clearer in larger-bodied organisms such as dinosaurs, we also see evidence for either competitive displacement or opportunistic replacement in smaller-bodied groups such as lepidosaurs, lissamphibians and mammaliaforms. There is some evidence that pterosaurs and paravian theropods rapidly diversified and adopted new ecomorphotypes in the Early Cretaceous, including the explosive radiation of the most successful extant tetrapod group, birds, although the precise timing of these events is obscured by varying spatiotemporal sampling of these clades. Low-latitude and shallow marine to semi-aquatic faunas, including testudines, crocodylomorphs, and reef-dwelling and sessile invertebrates, suffered the greatest diversity loss in the marine realm, whereas more mobile taxa with greater dispersal ability, such as ichthyosaurs, appear to have been relatively unaffected.[This next point is my favourite!]
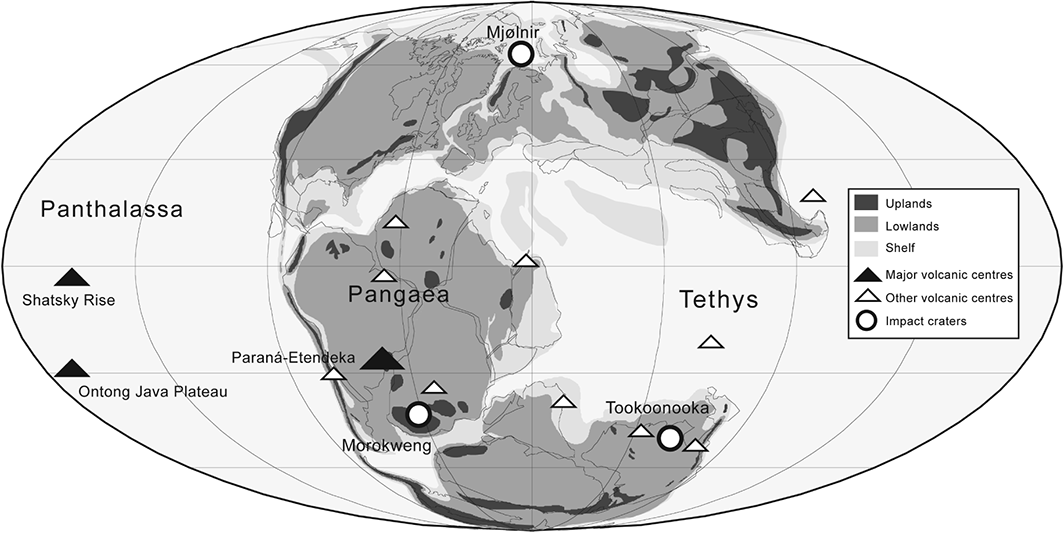
- The Late Jurassic–Early Cretaceous was a period of major environmental perturbations that have largely been ignored or overlooked in historical analyses of Mesozoic diversity dynamics, in favour of more ‘exotic’ extinction intervals. A range of evidence indicates the following major changes: (i) at least three large bolide impacts in the latest Jurassic, one of which might have been bigger than the end-Cretaceous Chicxulub impact (Fig. 3); (ii) a Late Jurassic–Cretaceous ‘greenhouse’ world, interrupted by a latest Jurassic ‘cold snap’ and corresponding aridity episode; (iii) a global drop in sea level to a eustatic lowstand through the J/K boundary (Fig. 2); (iv) potentially heightened levels of anoxia, oceanic stagnation, and sulphur toxicity over the J/K boundary; (v) a series of repeated ‘biocalcification crises’ in the Early Cretaceous, along with two purported oceanic anoxic events in the Valanginian and Hauterivian; (vi) the emplacement of the Paraná and Etendeka (late Valanginian–Hauterivian) and Ontong Java Plateau (Barremian–early Aptian) flood basalts, the latter of which might have been three times as voluminous as the end-Cretaceous Deccan volcanism; and (vii) some of the largest volcanic episodes in the history of the Earth, following the emplacement of the Shatsky Rise supervolcano at the J/K boundary. This series of environmental perturbations warrants further investigation in the context of potential biotic effects throughout this time.
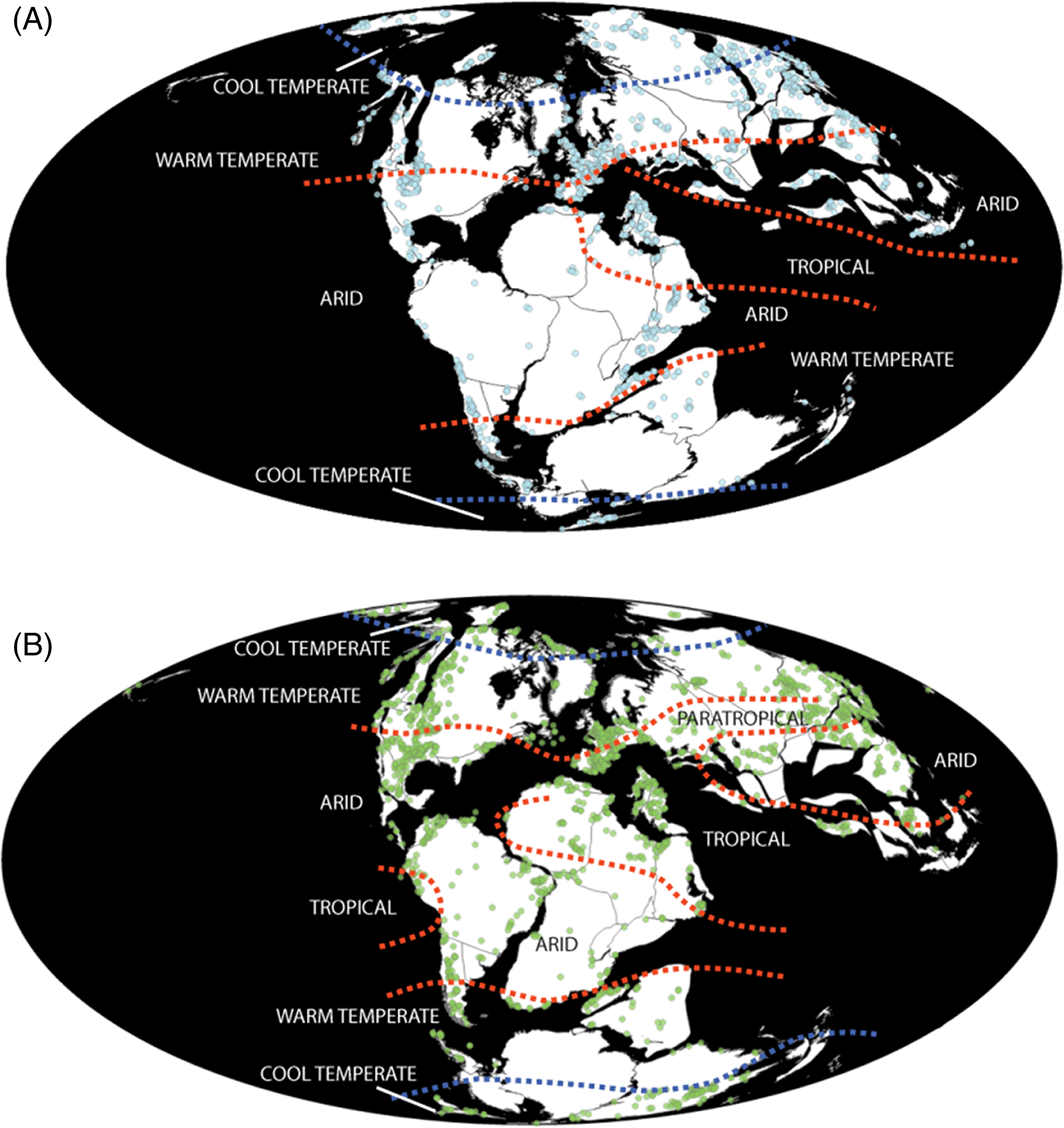
(A) Late Jurassic (Oxfordian–Tithonian) and (B) Early Cretaceous (Berriasian–Albian) global fossil occurrences, with climatic regions overlain. Fossil data extracted from The Paleobiology Database, November 2014 (http://paleobiodb.org/). Palaeoclimate data from Paleomap project (http://www.scotese.com/). (Source) - The J/K boundary represents an opportunity to investigate the environmental and ecological factors governing recovery (e.g. Wei et al., 2015). Distinct extinction and diversification patterns are clearly recorded in different groups, with a range of potential extrinsic abiotic controls. Additionally, the fact that a faunal turnover at the J/K boundary appears to be coupled with an ecological turnover in many groups, suggests that intrinsic biological parameters, principally regarding acquisition of key ecological characteristics and morphological plasticity and disparity, require further investigation in terms of the effects that these might have had on survivability. For example, low disparity in sauropterygians and turtles is coupled with strong evidence for a faunal turnover, whereas high ecological diversity in ichthyopterygians, lepidosaurs, and mammaliaforms is reflected in high survivability rates across the J/K boundary. This level of complexity necessitates the use of a multivariate approach to assessing macroevolutionary drivers (e.g. Benson & Mannion, 2012).
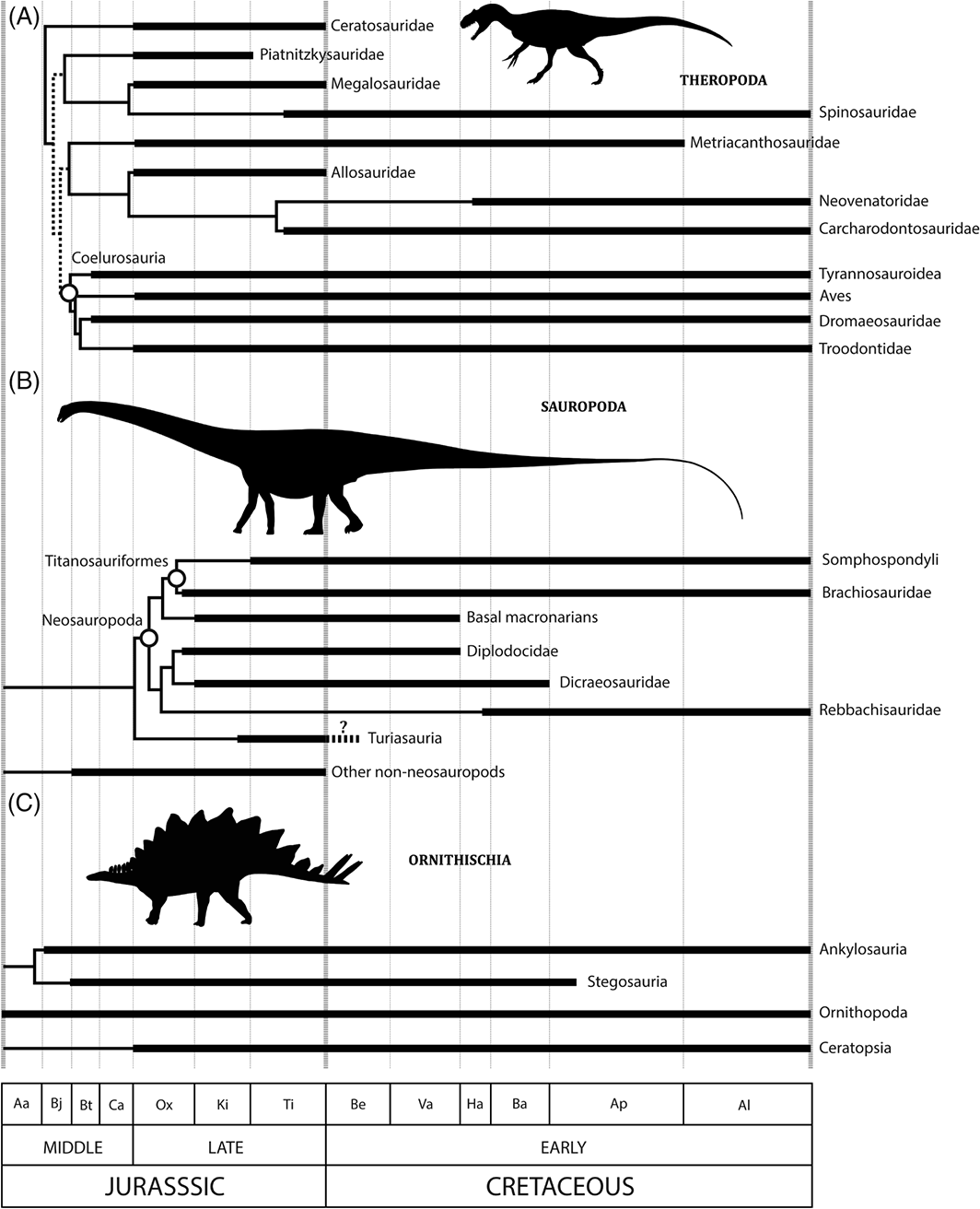
Stratigraphic ranges of major Jurassic–Cretaceous theropod (A), sauropod (B), and ornithischian (C) dinosaur clades through the Middle Jurassic to Early Cretaceous. (Source) - There are major gaps in our current knowledge of biological and Earth systems processes and patterns during the J/K interval. These include the absence of sampling-standardised diversity trajectories for many terrestrial and marine clades (e.g. plants, terrestrial insects, and small-bodied tetrapods), and the biotic and abiotic drivers of these patterns. Substantial progress has recently been made in modelling the possible drivers of diversification and extinction, especially in terrestrial tetrapods (Sookias, Benson & Butler, 2012a;Sookias, Butler & Benson, 2012b; Benson & Druckenmiller, 2014; Benson et al., 2014a,b) and marine invertebrates (e.g. Peters,2008). Combining these methods with increasingly sophisticated ways of analysing diversity in the fossil record (e.g. Alroy, 2010a,2014), will provide considerable insight into the macroevolutionary history of life through the J/K boundary.
So, there you have the short (ha) summary of the paper! I’ve got a couple of things in the works now which help to resolve some of these questions, and will be sure to post them on here as soon as they’re out!
Nice work Jon. I’m a big fan of volcanic eruptions these days. Oh that pesky Shatsky Rise!
What does the invertebrate record say/reveal? Was there as dramatic a turnover in the oceans as well? If not…
From the paper (Discussion section):
Articulate brachiopods, gastropods, bivalves and ammonites all declined in diversity at the J/K boundary, with the latter two groups showing evidence for regional selectivity (Alroy, 2010a; Rogov et al., 2010), alongside a higher extinction intensity in northern hemisphere taxa (Alroy, 2010b). Additionally, these groups exhibited latitudinal constraints on diversity, possibly driven by large-scale changes in global climate regimes at the J/K boundary (Anderson et al., 1999; Scotese et al., 1999; Bergman et al., 2004; Meyers, 2014). Such constraints might be responsible for global declines in diversity (Peters & Foote, 2001; Smith & McGowan, 2007; McGowan & Smith, 2008; Lu et al., 2009; Alroy, 2010b; Rogov et al., 2010; Valentine & Jablonski, 2010; Smith et al., 2012), and the reorganisation of marine ecosystems through the J/K boundary. Additional evidence suggests that climate strongly influenced oceanic productivity and nutrient cycles at the J/K boundary (Danelian & Johnson, 2001), which, combined with a eustatic lowstand (Miller et al., 2005), would have strongly impacted upon marine life. This could have provided a mechanism for the different patterns exhibited by shallow- and deep-water invertebrate taxa. There is a growing body of evidence that low-latitude reef-dwelling or shallow-marine and sessile epifauna (e.g. cemented bivalves, corals) were the most severely affected at the J/K boundary (Zakharov & Yanine, 1975; Skelton et al., 1990; Aberhan et al., 2006; Kiessling, 2008, 2009; Alroy, 2010a; Ruban, 2011; Foote, 2014), possibly due to a dramatic shift from calcitic to aragonitic organisms (Kiessling et al., 2008).
The decline of reefs over the J/K boundary was probably also tied to changes in global temperatures (Anderson et al., 1999; Scotese et al., 1999; Bergman et al., 2004; Martin-Garin et al., 2010). Additionally, it is likely that factors relating to sea-level changes, including declining salinity and shifts in nutrient flux systems, constrained organisms to increasingly rare shallower shelf systems over the J/K boundary and until the middle Cretaceous (Hay et al., 2006). The core driver for these changes in sea level and marine productivity (Danelian & Johnson, 2001) potentially relates to the connection between the Atlantic and the Pacific (Panthalassa) oceans during the J/K interval, with shorter term variation driven by fluctuations in the extent of polar ice caps (Haq, 2014). Large-scale tectonic processes at this time, particularly regarding the break-up of Pangaea, must have been important in controlling the biogeography of marine and terrestrial taxa through the latest Jurassic to earliest Cretaceous (Galton, 1982; Scotese et al., 1988; Scotese, 1991; Pérez-Moreno et al., 1999; Mateus, 2006; Escaso et al., 2007). However, whether sea level is alone in driving these documented extinctions, or whether dramatic climatic changes played a role (Anderson et al., 1999; Scotese et al., 1999; Bergman et al., 2004; Meyers, 2014) is presently unknown.
The J/K boundary further exhibits a dramatic decline in reef communities, illustrated by a distinct turnover from rudist to scleractinian-dominated systems (Aberhan et al., 2006). This reef decline might also be related to the dramatic rise in shallow to moderately deep infaunal suspension feeders in shallow marine settings (Aberhan et al., 2012). Congruent with this is the dramatic decline of other reef-dwelling epifauna such as crustaceans (Klompmaker et al., 2013), prior to a faunal turnover in brachyuran decapods that reflects an ecological shift to deeper-water taxa. However, some reef-dwelling organisms appear to have been comparatively unaffected (e.g. echinoderms), perhaps facilitated by a high ecological diversity in these groups (e.g. Baumeister & Leinfelder, 1998). In the majority of other marine invertebrate groups, there is less evidence for a faunal turnover, with the J/K boundary instead representing a time of elevated extinction rates, but not accompanied by within-group ecological reorganisation. These changes, however, are also likely to have been tied to the environmental changes outlined in Section II (b; therefore any attempt to decouple biotic and abiotic factors influencing marine diversity patterns remains problematic.
Thanks, very interesting and useful! At participating in MOOCs on Earth history [astrobiology interest] the faunal changes after the J/K boundary came over as something of a mystery.
Jon;
Protracted multiphase extinction over 25 Ma; totally with you on that. Have you had a chance to read the Cedar Mt. tome yet? https://www.utahgeology.org/openjournal/index.php/GIW/article/view/9
For North America
Clearly big faunal turnover across J-K boundary but still an appreciable gap of maybe 10 million years.
Much wetter climatic conditions seem to span the unconformity (link?) before the rainshadow formed by the Sevier orogeny.
There seems to be a big faunal break sometime in the medial Aptian. North America maybe larger than elsewhere as it becomes a island continent for the remainder of the Early Cret. before first Asian immigration event, with immigration induced extinction,
Ontong Java Plateau (more early Aptian) flood basalts??
More data coming relative to mammals, sauropods, and stegosaurs.
Enjoy, my friend!
Hey Jim,
Ah this is awesome! I hadn’t read it yet, thanks for sharing – I’ll get on it! I reckon it would be really cool to do a comprehensive review of the biotic/abiotic things happening in the Early Cretaceous of North America, seems like there’s so much potentially interesting things going on!
Cheers,
Jon
You should be interested by https://php5.univ-brest.fr/conference/ocs/index.php/JK2018/JK2018
protohedgehog,
I am not a scientist but have read about science since very young. I have long contemplated over the past several decades on possible causes during the KT extinction event that brought about the complete extinction of the land based dinosaurs with the exception of the birds that nested in the trees or on high cliffs. I was just reading this very interesting paragraph by Bob Strauss dated February 18, 2018 on the internet titled “Prehistoric Life During the Mesozoic” that conformed with some of my speculations:
“During most of the Mesozoic Era, including the Cretaceous period, mammals were sufficiently intimidated by their dinosaur cousins that they spent most of their time high up in trees or huddling together in underground burrows. Even so, some mammals had enough breathing room, ecologically speaking, to allow them to evolve to respectable sizes. One example was the 20-pound Repenomamus, which actually ate baby dinosaurs!”
I have read about certain sea birds who nest on the ground and have done so throughout most of their evolutionary past on secluded rodent and pest free islands. After the introduction of rodents to their islands by humans, it has been documented that the nesting birds are totally defenseless and seem to be blind to the rodents as the rodents literally chew and eat them and their young while they are sitting and tending their nests. There are photos of the nesting birds with large chunks of flesh eaten out of them while they sit on their nests. They seem to have evolved no defenses to the rodent predations.
Could it be that mammals like Repenomamus evolved to eat the eggs and hatchlings of the land based dinosaurs while the parenting dinosaurs looked on with indifference like today’s sea birds? And that the evolutionary niche filled by Repenomamus like mammals was so unchallenged that they multiplied by the billions and helped to push all land based dinosaurs into extinction with the exception of the birds who nested in the trees and high up on steep cliffs that may have been inaccessible to Repenomamus and any other dinosaur eating mammals? Is it possible that no land based dinosaurs made it through the KT extinctions because what dinosaurs the KT extinctions would have normally missed were finished off by mammals? It would be ironic that the small mammals of the cretaceous helped bring down their mighty dinosaur cousins that helped pave the way for the new age of mammals.
Small comment. Being more precise about siting the boundary, am not so sure significant extinctions can be detected – certainly there is nothing like any of the major Phanerozoic extinctions . In marine settings. just before the boundary there was a radiation of calcareous nannofossils and a marked turnover of calpionellids actually marks the J/K boundary (the Tithonian/Berriasian boundary): that is just above the old boundary level that was previously used – a level marked by a turnover in ammonite faunas (base of Jacobi Zone). My point is this – before hunting for an extinction event at the J/K boundary, to able to identify any faunal or floral event or extinction, it is important to remember that the base of the Cretaceous is the base of the Berriasian Stage, and that is at the base of the Calpionella alpina Zone. And none of this can be readily identified in non-marine settings.
Hi Bill, thanks so much for commenting! And yes, you’re totally right. In the paper this little post is drawn from, we addressed this issue a bit (based on your own work) (https://onlinelibrary.wiley.com/doi/10.1111/brv.12255) and also in the follow up too (https://www.nature.com/articles/ncomms12737) – I think most discussion is in the SI for this one. These issues are why we elected to use the term J/K transition mostly instead of J/K boundary itself. Certainly made this PhD tougher! 🙂
Jon, Thanks very much.
It is a difficult time period, with closing seaways, isolated basins, endemic invertebrates, discontinuous sedimentation and missing intervals on the continents. ‘Transition’ maybe helps. It is really a solid line drawn on the chronostratigraphic chart, as all stage boundaries are. Part of the problem with vertebrates (land or marine) between the Jurassic and Cretaceous is data, or the lack of it. One sees publications talking about evolutionary happenings at the boundary or across the boundary, when most of the fossil evidence actually comes from some way down in the Jurassic or well up into the Cretaceous – nowhere near the boundary. There are not so many places where non-marine rocks from Tithonian/Berriasian times survive – the Purbeck Formation of England is such a rarity – and deeper-water marine sequences seldom have vertebrates. There is much research in progress on the calibration between non-marine (Purbeck) and (more numerous) marine sequences using magnetostratigraphy. And, no doubt, more missing pieces of the faunal jigsaw will be found. We can be sure that the J/K interval and its faunas will remain a fascinating puzzle for years to come.
Wonderful explanation, particularly the faunal and environmental possible influences on the 25 million years of the transition period.
The question I would ask, is what is the floral evidence if any?